Ածխաջուր
Ածխաջրեր, (ածխաջրատներ, շաքարներ), քիմիական միացություններ՝ կազմված ածխածին, թթվածին և ջրածին տարրերից։ Ածխաջուր են կոչվում, որովհետև միացության մեջ ջրածին և թթվածին տարրերը գտնվում են ջրի մոլեկուլում ունեցած համամասնությամբ՝ Cx(H2O)y։ Կառուցվածքով և քիմիական հատկություններով ունեն շաքարների բնույթ։ Սպիտակուցների և ճարպերի հետ միասին ածխաջրերը կարևոր նշանակություն ունեն մարդու և կենդանիների օրգանիզմներում ընթացող նյութերի ու էներգիայի փոխանակության շարժընթացում։ Մտնում են բուսական, կենդանական և բակտերային օրգանիզմների կազմության մեջ։ Ածխաջրերը մարդու և կենդանիների սննդի կարևոր բաղադրամաս են և ապահովում են դրանց կենսագործունեության համար անհրաժեշտ էներգիան։ Հասուն մարդու օրգանիզմում էներգիայի կեսից ավելին առաջանում է ածխաջրերից[1]։

Նշանակություն
խմբագրել
Ածխաջրերը կարևոր նշանակություն ունեն որպես հիմնական շինանյութ բույսերի և կմախք՝ միջատների, խեցգետնակերպերի ու մյուս օրգանիզմների համար։ Մտնում են բջջապատի, շարակցական հյուսվածքի հիմնական նյութի կազմության մեջ, բացի այդ, բարդ կենսապոլիմերների, օրինակ՝ գլիկոպրոտեիդների բաղադրության մեջ ածխաջրերը կարող են լինել կենսաբանական ինֆորմացիայի կրողներ և պայմանավորել այդ միացությունների իմունաբանական առանձնահատկությունը։ Այսպես՝ արյան պատկանելությունը որևէ խմբի որոշվում է արյան խմբային նյութերի կազմության մեջ մտնող ածխաջրերի կառուցվածքով և հաջորդականությամբ։ Հաստատվել է բջիջների մակերևույթի նյութերի բաղադրիչ մասը կազմող ածխաջրերի կարևոր նշանակությունը բջիջների փոխներգործություններում։ Նույն տիպի բջիջների միմյանց «ճանաչումը», տարբերակումը, աճումը, կենսապոլիմերների (սպիտակուցներ, նուկլեինաթթուներ և այլն) արտազատումը պայմանավորված են բջիջների մակերևույթի ածխաջրերի առանձնահատուկ գործունեությամբ։ Պարզվել է, որ դրանք կարևոր են չարորակ ուռուցքների առաջացման, վիրուսների ու բջիջների փոխներգործության գործընթացներում։ Մարդու և կենդանիների օրգանիզմում որոշ բարդ ածխաջրեր (օրինակ՝ հիալուրոնաթթուն) «քսանյութերի» ֆունկցիա են կատարում և ծառայում են որպես հեղուկ միջավայր բջիջների տեղաշարժման համար, դրանցով պատվում են շփվող, օրինակ՝ հոդային մակերևույթները։ Որոշ ածխաջրեր (վիտամին C, վիտամին B15, հեպարին) օժտված են առանձնահատուկ կենսաբանական ակտիվությամբ[2]։
Էներգիական նշանակություն
խմբագրելԷներգիական արժեքով ածխաջրերը հավասարարժեք են սպիտակուցներին։ 1 գ ածխաջրի այրումից օրգանիզմում անջատվում է միջինը 4,1 կկալ էներգիա։ Մարդու սննդի մեջ ածխաջրերի քանակը սովորաբար 4 անգամ գերազանցում է սպիտակուցների և ճարպերի պարունակությունը։ Օրգանիզմում ածխաջրերի սահմանափակ կուտակվելու հատկությունը նպաստում է դրանց ավելցուկային քանակների հեշտորեն ճարպի փոխարկվելուն, որը կուտակվում է ճարպային պահեստարանում։ Սննդի մեջ ածխաջրերի ավելցուկից առաջանում է ավելորդ քաշ և գիրություն։ Ուստի պետք է խուսափել նաև ածխաջրերի չարաշահումից։
Ածխաջրերը սննդում անհրաժեշտ են այնպես, ինչպես սպիտակուցները և լիպիդները։ Ածխաջրերի անհրաժեշտությունը բնորոշվում ոչ միայն նրանով, որ նրանք հանդիսանում են «արագընթաց» էներգիայի աղբյուր առանց կողմնակի թունավոր արգասիքների, այլ նաև նրանց մասնակցությամբ կենդանի բջջի բարդ սպիտակուցների՝ գլիկոպրոտեիդների կարգավորիչ և պաշտպանական ֆունկցիաներին։ Լիպիդների օքսիդացումը ածխաջրային էներգիայի պակասի դեպքում ծայրահեղ դանդաղ է ընթանում[3]։
Էներգիայի աղբյուր են հանդիսանում հեշտ յուրացվող ածխաջրերը՝ գլյուկոզ, ֆրուկտոզ, սախարոզ, օսլա, գլիկոգեն։ Այդ ածխաջրերի փոխանակությունը մարդու մոտ սերտորեն կապված է լիպիդների և ճարպերի փոխանակության հետ։ Ածխաջրերը սննդի հետ ավելցուկով ընդունելու դեպքում հեշտությամբ փոխարկվում են ճարպերի, բայց լիպիդներից ածխաջրեր գործնականում չեն սինթեզվում[4]։
Ածխաջրային էներգիայի պակասը առողջ մարդու մոտ կոմպենսացվում է նրանց սինթեզի ակտիվացմամբ՝ ամինաթթուների փոխանակության միջանկյալ արդյունքների, մակերիկամների հորմոնների և որոշ վիտամինների, այդ թվում C վիտամինի մասնակցությամբ։ Այդ պրոցեսը կոչվում է գլյուկոնոգենեզ։ Այդ պատճառով որոշակի սահմաններում սննդի սպիտակուցա-լիպիդային ուղղվածության դեպքում, կարելի է սահմանափակել ածխաջրերով հարուստ մթերքի օգտագործումը առանց օրգանիզմին վնասելու։
Արդյունաբերական նշանակություն
խմբագրելԱծխաջրերն ունեն նաև արդյունաբերական նշանակություն. օսլան օգտագործվում է գլյուկոզ և էթիլսպիրտ ստանալու համար, ինչպես նաև թղթի ու մանածագործական արդյունաբերության մեջ։ Օսլան մտնում է դեղանյութերի, քսուքների, լաքերի բաղադրության մեջ, նրանից պատրաստում են սոսինձ, դեղապատիճներ և այլն։ Փայտանյութից ստացված թաղանթանյութն օգտագործվում է թուղթ, արհեստական մետաքս, պայթուցիկ նյութեր պատրաստելու համար։
Մոնոսախարիդներ
խմբագրելՄոնոսախարիդները ըստ կառուցվածքի պոլիհիդրօքսիկարբոնիլային միացություններ են (պոլիհիդրօքսիալդեհիդներ և պոլիհիդրօքսիկետոններ)։ Նրանք պինդ, ջրում լավ լուծելի, հիմնականում քաղցրահամ, միացություններ են։ Ջրային լուծույթներն ունեն չեզոք ռեակցիա։
Մոնոսախարիդների դասակարգումը
խմբագրելՄոնոսախարիդների դասակարգումը հիմնված է կառուցվածքային յուրահատկությունների վրա։ Ըստ կարբոնիլային խմբի բնույթի տարբերում են ալդոզներ (-CHO) և կետոզներ (-C=O),իսկ շղթայում ածխածնի ատոմների թվի՝ տրիոզներ, տետրոզներ, պենտոզներ, հեքսոզներ և հեպտոզներ։
Կենսաբանական համակարգերում կարևոր նշանակություն ունեն պենտոզները և հեքսոզները, որոնք, հաշվի առնելով վերը նշված հատկանիշներից, բաժանվում են ալդոպենտոզների, ալդոհեքսոզների, կետոպենտոզների, կետոհեքսոզների։ Քանի որ յուրաքանչյուր մոնոսախարիդի կազմում կա մի քանի ասիմետրիկ ածխածնի ատոմ, հետևաբար մոնոսախարիդները առաջացնում են տարբեր ստերեոիզոմերներ (օրինակ, գլյուկոզ, մանոզ, գալակտոզ՝ բոլորը ալդոհեքսոզ են, մտնում են ալդոզների ընտանիքի մեջ և տարբերվում են իրենց ֆիզիկական հատկություններով)։
| Ալդոտրիոզներ |  գլիցերալդեհիդ (գլիցերալ) | |||||||
| Ալդոտետրոզներ |  D-էրիթրոզա |
 D-տրեոզա | ||||||
| Ալդոպենտոզներ |  D-ռիբոզա |
 D-արաբինոզա |
 D-քսիլոզա |
 D-լիկսոզա | ||||
| Ալդոհեքսոզներ |  D-ալլոզա |
 D-ալտրոզա |
 D-գլյուկոզա |
 D-մաննոզա |
 D-գուլոզա |
 D-իդոզա |
 D-գալակտոզա |
 D-տալոզա |
| Կետոտրիոզներ | Դիհիդրոքսիացետոն | |||
| կետոտետրոզներ |  էրիտրուլոզա | |||
| կետոպենտոզներ |  ռիբուլոզա |
 քսիլուլոզա | ||
| կետոհեքսոզներ |  պսիկոզա |
 ֆրուկտոզա |
 սորբոզա |
 տագատոզա |

Մոնոսախարիդների ստերեոիզոմերների թիվը որոշվում է 2n բանաձևով, որտեղ n-ասիմետրիկ ածխածինների թիվն է։ Ալդոպենտոզների ստերեոիզոմերների թիվը հավասար է 8 (23),և նրանք կազմում են չորս զույգ էնանտիոմերներ։ Ալդոհեքսոզների ստերեոիզոմերների թիվը հավասար է 16 (24) է, նրանք կազմում են 8 զույգ էնանտիոմերներ։ Մոնոսախարիդները պատկանում են երկու ստերեոիզոմերների շարքի՝ D և L: Ածխաջրերի պարզագույն ներկայացուցիչներն են ալդե- և կետոտրիոզները։

D և L շարքերի պատկանելիությունը որոշվում է ամենամեծ համարը կրող ասիմետրիկ ածխածնի կոնֆիգուրացիայով, այն համեմատելով D կամ L- գլիցերալդեհիդի կոնֆիգուրացիայի հետ։ Տվյալ մոնոշաքարի անվանումը որոշվում է բոլոր խիրալ կենտրոնների կառուցվածքով[5]։


Մոնոսախարիդները օպտիկապես ակտիվ միացություններ են.նրանց լուծույթները պտտում են բևեռացված լույսի հարթությունը։ Բևեռացված լույսի հարթության պտտման մեծությունը և ուղղությունը յուրահատուկ է յուրաքանչյուր ստերեոիզոմերի համար, կապված չէ D կամ L շարքերի պատկանելիության հետ և որոշվում է փորձնական ճանապարհով։ Բնական ծագում ունեցող մոնոսախարիդները հիմնականում D-շարքի ներկայացուցիչներն են[6]։
Մոնոսախարիդների կառուցվածքի ընդունված է պատկերել Ֆիշերի պրոյեկցիոն բանաձևերով, որոնցում ասիմետրիկ ածխածինը չի նշվում։
Մոնոսախարիդների ցիկլիկ կառուցվածքը
խմբագրել
Մոնոսախարիդները ցուցաբերում են բոլոր այն հատկությունները, որոնք պայմանավորված են նրանց կազմի մեջ մտնող ֆունկցիոնալ խմբերով։ Սակայն որոշ քիմիական հատկություններ հնարավոր չէ բացատրել ելնելով վերը բերված կառուցվածքից։
-
 β-D-ֆրուկտոֆուրանոզ
β-D-ֆրուկտոֆուրանոզ -
 α-D-ռիբոֆուրանոզ
α-D-ռիբոֆուրանոզ -
 β-ռիբոֆուրանոզ
β-ռիբոֆուրանոզ -
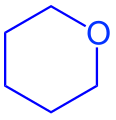 Պիրանոզ կառուցվածք Հեուորսի բանաձևով
Պիրանոզ կառուցվածք Հեուորսի բանաձևով




Առաջինը ռուս գիտնական Ա.Ա. Կոլինը կարծիք հայտնեց այն մասին, որ մոնոսախարիդներն ունեն ցիկլիկ կառուցվածք։ Լինելով հիդրօքսիալդեհիդներ (կամ կետոններ),մոնոսախարիդների բաց ձևերը ինքնաբերաբար կարող են վերածվել ցիկլիկ կառուցվածք ունեցող կիսաացետալների։ Օրինակ, ռիբոզի ճանկաձև (աքցանաձև) կոնֆիգուրացիայի կարբոնիլային խումբը և 4-րդ կամ 5-րդ ածխածնի OH խումբը տարածության մեջ մոտ են գտնվում և OH-ի թթվածնի նուկլեաֆիլ ատոմը գրոհում է կարբոնիլային ածխածնի էլեկտրաֆիլ ատոմի վրա։ Առաջանում է ցիկլիկ կիսաացետալ։ Կախված ցիկլիկ չափսերից, առաջացող միացությունները կոչվում են ֆուրանոզային (հինգանդամանի) կամ պիրանոզային (վեցանդամանի)։
Ցիկլիկ ձևում պրոխիրալ (կարբոնիլային խմբի ածխածինը) ոչ սիմետրիկ կենտրոնը վերածվում է խիրալ կենտրոնի։ Հետևաբար աճում է ստերեոիզոմերների թիվը։ Առաջացած կիսաացետալային OH խումբը կոչվում է գլիկոզիդային, իսկ ՛՛C-1՛՛ ածխածինը՝ անոմերային։ Ըստ գլիկոզիդային խմբի դիրքի տարբերում են α- և β-անոմերներ։ Ներմոլեկուլային փոխազդեցության հետևանքով առաջանում են 5 և 6 անդամանի թերմոդինամիկորեն կայուն ցիկլեր՝ ֆուրանոզային և պիրանոզային։
Մոնոսախարիդների ցիկլիկ կառուցվածքները ընդունված է պատկերել նաև Հեուորսի բանաձևերով։
Հեուորսի բանաձևերում պիրանոզային ցիկլի թթվածինը պատկերում են աջ հեռավոր անկյունում (C-1-ը՝ աջից)։OH-խմները, որոնք բաց ձևերի մոտ (Ֆիշերի բանաձևերում) գտնվում էին աջ կողմից գրվում են ցիկլի հարթությունից ցածր, իսկ ձախ կողմից գտնվող OH-խմբերը՝ հարթությունից վերև։ Այն անոմերները, որոնցում գլիկոզիդային խումբը պատկերում են հարթության տակ, կոչվում են α-անոմերներ։ Անոմերները, որոնցում գլիկոզիդային խումբը պատկերում են հարթությունից վեր կոչվում են β-անոմերներ։
Ցիկլո-օքսո տաուտոմերիա
խմբագրելԱնոմերները տարբերվում են հալման, եռման ջերմաստիճաններով, օպտիկական հատկություններով։ Հայտնի է, որ α-D-գլյուկոպիրանոզի թարմ լուծույթի տեսակարար պտտման անկյունը հավասար է 112°, իսկ β-D-գլյուկոպիրանոզինը՝ 19°:
Մոնոսախարիդի բոլոր 5 ձևերը (ացիկլիկ, α- և β-ֆուրանոզային և α- և β պիրանոզային) լուծույթում գտնվում են դինամիկ հավասարակշռության մեջ։ Առավել կայուն են մոնոսախարիդների պիրանոզային ձևերը։
Օրինակ, գլյուկոզի ջրային լուծույթում 99,9% կազմում է գլյուկոզի պիրանոզային ձևը։ Բաց ձևը կազմում է ընդամենը 0,003-0,03% և միջանկյալ է՝ ցիկլիկ ձևերը փոխարկվում են մեկը մյուսին բաց ձևի միջոցով։ Այսինքն գոյություն ունեն տաուտոմերիայի ևս մեկ տեսակ՝ ցիկլո-օքսո։
Դրանով է բացատրվում այն փաստը, որ բյուրեղային α- և β-D-գլյուկոպիրանոզներից պատրաստված թարմ լուծույթների բևեռացված լույսի պտտման անկյունը, որը համապատասխանաբար հավասար է +112° և +19°, մի քանի ժամ հետո փոխվում է և հավասարվում +52,7°: Դա պայմանավորված է նրանով, որ լուծույթում հաստատվում է դինամիկ հավասարակշռություն տարբեր իզոմերների միջև, որում գերակշռում են α- և β-պիրանոզային ձևերը (1:2 հարաբերությամբ, 36% α- և 64% β-)։ Մոնոսախարիդների թարմ լուծույթների ժամանակի ընթացքում բևեռացված լույսի պտտման անկյան փոփոխության երևույթը կոչվում է մուտառոտացիա։ Այդ պատճառով լուծույթում գտնվող մոնոսախարիդի համար սովորաբար չեն նշվում անոմերային ածխածնի կոնֆիգուրացիան։
-
β-D-ֆրուկտոֆուրանոզ
-
 α-D-ֆրուկտոֆուրանոզ
α-D-ֆրուկտոֆուրանոզ

Մոնոսախարիդների կոնֆորմացիաները
խմբագրելՔանի որ վեցանդամանի (պիրանոզային) ցիկլը հարթ չէ, Հեուորսի բանաձևերը լրիվ պատկերացում չեն տալիս մոնոսախարիդների ցիկլիկ ձևի տարածական կառուցվածքի մասին։ Ինչպես և ցիկլոհեքսանի դեպքում, ածխաջրերի առավել կայուն կոնֆորմացիան բազկաթոռն է։

 Կոնֆորմացիոն կառուցվածքով է բացատրվում D-գլյուկոզի առավել կայուն լինելը՝ նրանում առաջնային սպիրտային և բոլոր հիդրօքսիլ խմբերը (բացառությամբ կիսաացետալային խմբի, այն էլ միայն α-անոմերի դեպքում) գտնվում են էկվատորիալ դիրքում։ Յուրաքանչյուր այլ հեքսոզ ունի գոնե մեկ աքսիալ OH-խումբ։
Կոնֆորմացիոն կառուցվածքով է բացատրվում D-գլյուկոզի առավել կայուն լինելը՝ նրանում առաջնային սպիրտային և բոլոր հիդրօքսիլ խմբերը (բացառությամբ կիսաացետալային խմբի, այն էլ միայն α-անոմերի դեպքում) գտնվում են էկվատորիալ դիրքում։ Յուրաքանչյուր այլ հեքսոզ ունի գոնե մեկ աքսիալ OH-խումբ։
Կոնֆորմացիոն կառուցվածքների հարաբերական կայունությամբ է բացատրվում գլյուկոզի α- և β-պիրանոզային ձևերի քանակները լուծույթում։ Տաուտոմեր հավասարակշռությունը շեղված է դեպի β-D-անոմերի առաջացում, քանի որ առավել կայուն են տեղակալիչների էկվատորիալ դասավորություն ունեցող կոնֆորմերները։

Մոնոսախարիդի կոնֆիգուրացիան, հատկապես անոմերային ածխածնինը (α- կամ β-) չափազանց կարևոր է պոլիսախարիդների տարածական կառուցվածքի ձևավորման համար, էկվատորիալ դիրքերի դեպքում առաջացած շղթան ունի գծային կառուցվածք (ցելյուլոզ),իկս աքսիալ դասավորվածության դեպքում առաջանում են գալարներ և ձևավորվում է պարուրաձև երկրորդային կառուցվածք (օսլա)։

Մոնոսախարիդների քիմիական հատկությունները
խմբագրելՄոնոսախարիդները, շնորհիվ հետերոֆունկցիոնալ կառուցվածքի (բազմաթիվ ռեակցիոն կենտրոնների առկայության) չափազանց ռեակցիոնունակ են։ Կիսաացետալային հիդրօքսիլով են պայմանավորված մոնոսախարիդների կարևորագույն հատկությունները։ Մոնոսախարիդները չոր HCl-ի ներկայությամբ սպիրտների հետ փոխազդելիս (անջուր պայմաններում),առաջացնում են ացետալներ, որոնք կոչվում են գլիկոզիդներ՝ կիսաացետալային խումբը փոխարինվում է նուկլեաֆիլ տեղակալման մեխանիզմով։ Առաջանում է α- և β- գլիկոզիդների խառնուրդ։

Անոմերային գլիկոզիդների խառնուրդը կարելի է պատկերել մեկ բանաձևով։
Մոնոսախարիդները գլիկոզիդներ են առաջացնում նաև մոնոսախարիդների սպիրտային խմբերի հաշվին, այսինքն առաջանում են օլիգո- և պոլիսախարիդներ։ Բացի այդ, օրգանիզմում տեղի է ունենում D-գլյուկուրոնաթթվի գլիկոզիդների առաջացում՝ որոշ տոքսիկ ֆենոլային միացությունների հետ։
Այդ գլիկոզիդները (գլյուկուրոնիդները) հեռանում են մեզով և տեղի է ունենում դետոքիկացում։O-գլիկոզիդները նման են ազոտ պարունակող անալոգները՝ N-գլիկոզիդները, որոնք հանդես են գալիս որպես նուկլեոտիդների, նուկլեինաթթուների բաղադրամասեր։
Օրգանիզմում գլիկոզիդների առաջացմանը մասնակցում են մոնոսախարիդների ֆոսֆատները կամ նուկլեոզիդդիֆոսֆատները։ Այս միացություններում ֆոսֆորական թթվի մնացորդը և նուկլեոզիդդիֆոսֆատը լավ հեռացող խմբեր են։
Գլիկոզիդները կարող են հիդրոլիզվել թթվային միջավայրում (ացետալների նման),բայց բավականին կայուն են նոսր հիմքերի հանդեպ։ Գլիկոզիդների հիդրոլիզը, ըստ էության, առաջացման ռեակցիայի հակառակն է՝ արդյունքում առաջանում է սպիրտ և մոնոսախարիդ։ Օրինակ, մեթիլ-β-D-գլյուկոպիրանոզիդի հիդրոլիզի արդյունքում առաջանում են գլյուկոզ և մեթանոլ։
Սպիրտային OH-խմբերի մասնակցությամբ ընթացող ռեակցիաներ.պարզ և բարդ եթերների առաջացում։
Պարզ եթերներ առաջանում են հիմնային միջավայրում գլիկոզիդների (կամ մոնոսախարիդների) և ալկիլհալոգենիդների (կամ այլ ալկիլացնող ագենտների) փոխազդեցության արդյունքում առաջացած գլիկոզիդների եթերների թթվային հիդրոլիզով։ Օրինակ, մեթիլ-α-D-գլյուկոպիրանոզի ալկիլացման արդյունքում առաջանում է մեթիլտետրա-օ-մեթիլ-α-D-գլյուկոպիրանոզիդ։
Առաջացած մեթիլտետրա-օ-մեթիլ-α-D-գլյուկոպիրանոզիդը ենթարկվում է թթվային հիդրոլիզի, վերածվելով տետրա-օ-մեթիլ-α-D-գլյուկոպիրանոզի՝ եթերի։ Մոնոսախարիդների ալկիլացման ընթացքում նույնպես առաջանում են գլիկոզիդների եթերներ, որոնց գլիկոզիդային կապը թթվային միջավայրում ենթարկվում է հիդրոլիզի և առացանում է պարզ եթեր։
Բարդ եթերները առաջանում են մոնոսախարիդների և ացիլացնող ագենտների, օրինակ՝ կարբոնաթթուների անհիդրիդների, փոխազդեցության դեպքում։
Մոնոսախարիդների փոխարկումներում մեծ դեր ունեն ֆոսֆատները (ֆոսֆորական թթվի բարդ եթերները)։ Բացի արդեն հիշատակված գլյուկոզո-1-ֆոսֆատից, կարևոր նշանակություն ունեն գլյուկոզո-6-ֆոսֆատը, ֆրուկտոզո-6-ֆոսֆատը, դիֆոսֆատը, օրինակ՝ ֆրուկտոզ-1,6-դիֆոսֆատը (ֆրուկտոզ-1,6-բիֆոսֆատ)։
Մոնոսախարիդների էսթերները, ինչպես և բոլոր բարդ եթերները, ընդունակ են հիդրոլիզվելու թթվային և հիմնային միջավայրերում մինչև համապատասխան թթու (աղ՝ հիմնայինի դեպքում) և սպիրտ (տվյալ դեպքում մոնոսախարիդը սպիրտ է)։
Մոնոսախարիդների օքսիդացման ռեակցիաները։ Ալդոզների օքսիդացումը չեզոք և թթվային միջավայրերում ընթանում է առանց մոլեկուլների ածխածնային շղթայի փոփոխության։ Կախված օքսիդացման պայմաններից, կարող են օքսիդանալ միայն ալդեհիդային խումբը, ալդհեհիդային և սպիրտային խմբերը և այլն, և արդյունքում առաջանում են տարբեր միացություններ։
Օրինակ, գլյուկոզի օքսիդացման ժամանակ առաջանում են գլյուկոնաթթու, գլյուկուրոնաթթու, գլյուկարաթթու։
Մեղմ օքսիդիչի դեպքում (բրոմաջուր) օքսիդանում է միայն ալդեհիդային խումբը և ռեակցիայի արդյունքում առաջանում են ալդոնաթթուների, գլյուկոզի դեպքում՝ գլյուկոնաթթու։
Կենդանի օրգանիզմում նման օքսիդացումը ընթանում է համապատասխան ֆերմենտի մասնակցությամբ։
Սպիրտները, ինչպես հայտնի է, օքսիդանում են առաջացնելով կարբոնաթթուներ, ըստ որում առաջնային սպիրները երկրորդայինների հետ համեմատ օքսիդանում են ավելի արագ։
Ուժեղ օքսիդիչի ազդեցության ժամանակ (նոսր ազոտական թթու) օքսիդանում են և՛ ալդեհիդային, և՛ առաջնային սպիրտային խմբերը, առաջացնելով գլիկարաթթուներ կամ շաքարաթթուներ։
Այսպես, գլյուկոզի օքսիդացումից առաջանում է գլյուկարաթթու, մանոզի-մանարաթթու, գալակտոզի-գալակտարաթթու։
Կենդանի օրգանիզմներում կարևոր նշանակություն ունեն գլյուկուրոնաթթուները, որոնք առաջանում են առաջնային սպիրտային խմբի օքսիդացումից։ Օրինակ, գլյուկուրոնաթթուն, որը բազմազան ֆունկցիաներ է կատարում, մասնակցում է թունազրկման ռեակցիաներին, որպես կառուցվածքային հատված մտնում է պոլիսախարիդների կազմի մեջ և այլն։
Մոնոսախարիդներից ուրոնաթթուների առաջացման դեպքում անհրաժեշտ է «պաշտպանել» ալդեհիդային խումբը.օրինակ, օքսիդացումից առաջ, մոնոսախարիդը վերածել գլիկոզիդի։ Մոնոսախարիդների գլիկոզիդների օքսիդացման արդյունքում առաջանում են գլիկուրոնաթթուների գլիկոզիդներ, որոնց թթվային հիդրոլիզից առաջանում են ազատ գլիկուրոնաթթուներ։
Օրգանիզմում այդ պաշտպանական դերը կատարում է 1-ֆոսֆատային խումբը։
Մոնոսախարիդների վերականգնման հետևանքով առաջանում են ալդիտներ (շաքարային սպիրտներ),որոնք քաղցրահամ են։ Վերականգնումը ընթանում է խիստ պայմաններում, ուժեղ վերականգնիչների մասնակցությամբ։
Ալդիտներից մի քանիսը, օրինակ, քսիլիտի (D-քսիլոզից) և սորբիտը (սորբիտոլ,D-գլյուկոզից) շաքարախտով հիվանդները օգտագործում են որպես շաքարի փոխարինիչ։ Ալդիտները չեն ընդգրկվում նյութափոխանակության մեջ, համապատասխան ֆերմենտների բացակայության պատճառով։
Ալդոզների և կետոզների փոխադարձ փոխարկումը։ Հիմնային միջավայրում տեղի է ունենում մոնոսախարիդների իզոմերացում։CH-կենտրոնի առկայության շնորհիվ, հիմնային միջավայրում, տեղի է ունենում ալդոզներից կետոզների առաջացում (իզոմերացում) և էպիմերացում C-2 մոտ։
Օրգանիզմում նման փոխարկումները ընթանում են ֆերմենտների մասնակցությամբ։ Այս փոխարկումների նշանակությունը չափազանց մեծ է, քանի որ մոնոսախարիդներից կենդանական համակարգերում միայն գլյուկոզն ունի փոխանակման համար անհրաժեշտ ֆերմենտների համակարգ։ Հետևաբար, այլ մոնոշաքարները փոխանակվելու համար պիտի վերածվեն գլյուկոզի։ Նկարագրված եղանակով ֆրուկտոզից և մանոզից առաջանում է գլյուկոզ։
Մոնոսախարիդների հայտնաբերման ռեակցիաները
խմբագրելՊենտոզների և հեքսոզների տարբերիչ ռեակցիաները
խմբագրելՀանքային թթուների հետ տաքացնելիս մոնոսախարիդները ենթարկվում են դեհիդրատացման (կորցնում են երեք մոլեկուլ ջուր) և ալդոպենտոզներից առաջանում է ֆուրֆուրոլ, իսկ ալդո- և կետոհեքսոզներից՝ 5-հիդրօքսիմեթիլֆուրֆուրոլ։ Ֆուրֆուրոլը և 5-հիդրօքսիմեթիլֆուրֆուրոլը ֆենոլների և արոմատիկ ամինների հետ տալիս են կոնդեսացման ռեակցիաներ, առաջացնելով գունավոր միացություններ։ Այս ռեակցիաների վրա են հիմնված պենտոզների և հեքսոզների քանակական և որակական հայտնաբերման մի շարք եղանակներ։
Օրինակ, ֆուրֆուրոլը փոխազդելով անիլինի, իսկ 5-հիդրօքսիմեթիլֆուրֆուրոլը՝ ռեզորցինի հետ առաջացնում են կարմիր գույնի միացություններ։
Մոնոսախարիդների հայտնաբերման որակական ռեակցիաները
խմբագրելԱլդեհիդային (կարբոնիլային) խմբի օքսիդացման վրա են հիմնված մոնոսախարիդների հայտնաբերման (որակական) ռեակցիաները։ Չնայած ալդեհիդային (բաց) ձևի բաժինը տաուտոմեր խառնուրդում չափազանց փոքր է, բայց նա անընդհատ առաջանում է և մնում հաստատուն։ Տոլենսի (Ag+),Բենեդիկտի, Ֆելինգի (Cu2+) ռեակտիվների օգնությամբ կարելի է հայտնաբերել ալդոզները։ Հիմնային միջավայրում մետաղների իոնները վերականգնվում են, իսկ մոնոսախարիդները քայքայվում են՝ առաջացնելով օքսիդացման արդյունքների խառնուրդ։
RCHO+Ag(NH3)2+→Ag↓+օքսիդացման արդյունքներ
RCHO+Cu2+(կոմպլեքս)→Cu2O↓+օքսիդացման արդյունքներ
Այս ռեակտիվների հետ դրական ռեակցիա տվող մոնոսախարիդները և նրանց ածանցյալները կոչվում են վերականգնող շաքարներ։ Գլիկոզիդները չեն ցուցաբերում վերականգնիչ հատկություններ և չեն փոխազդում այս ռեագենտների հետ։ Սակայն, այս ռեակցիաները, ի տարբերությունյ բրոմաջրով և ազոտական թթվով օքսիդացման, չի կարելի օգտագործել ալդոնաթթուների ստացման համար, քանի որ հիմնային միջավայրում առաջանում է ենդիոլային ֆրագմենտ, և կարող է տեղի է ունենալ իզոմերացում (օրինակ գլյուկոզը վերածվի մանոզի կամ ֆրուկտոզի),ինչպես նաև ածխածնային շղթայի քայքայում։
Մոնոսախարիդների ածանցյալները
խմբագրելՄոնոսախարիդների ածանցյալներ են համարվում մոնոսախարիդային բնույթի այն միացությունները, որոնք մեկ կամ մի քանի հիդրօքսիլային խմբերի փոխարեն պարունակում են ջրածնի ատոմ կամ ֆունկցիոնալ խմբեր։ Օրինակ, նուկլեինաթթուների կազմի մեջ մտնում է 2-դեզօքսի-D-ռիբոզը, հետերոպոլիսախարիդների կազմի մեջ հանդիպում են մոնոսախարիդների ամինաածանցյալները։
Առավել կարևոր նշանակություն ունեն մոնոսախարիդների 2-ամինաածանցյալները, օրինակ,D-գլյուկոզամինը։
Բացի այդ, հանդիպում են նաև ամինախմբի ալկիլացված կամ ացիլացված ածանցյալներ՝ N-ացետիլգլյուկոզամին,N-ացետիլգալակտոզամին և այլն։
Նեյրամինաթթուն և նրա ածանցյալները, սիալաթթուները (N-ացետիլնեյրամինաթթուները) արյան և հյուսվածքների սպեցիֆիկ միացությունների բաղադրամասերն են։ Ազատ վիճակում գտնվում են ողնուղեղային հեղուկում։ Նեյրամինաթթվի սինթեզը կատարվում է պիրոխաղողաթթվից և մանոզամինից։
Ասկորբինաթթուն (վիտամին C) ըստ կառուցվածքի 2-կետո-L-գուլոնաթթվի γ-լակտոն է։
Ասկորբինաթթուն նյութափոխանակության ընթացքում մասնակցում է զանազան վերականգնման պրոցեսների։ Մասնավորապես կարևոր նշանակություն ունի կոլագենի կազմում պրոլինի հիդրօքսիլացմանը մասնակից ֆերմենտի (պրոլինհիդրօքսիլազի) գործունեության համար։ Ռեակցիայի արդյունքում ասկորբինաթթուն օքսիդանում է, վերածվելով դեհիդրոասկորբինաթթվի։
Արդյունաբերական եղանակներով ասկորբինաթթուն մեծ քանակներով ստանում են գլյուկոզից։ Վիտամին C-ի անբավարարության դեպքում զարգանում են զանազան հիվանդություններ (սկորբունտ, լնդախտ)։
Օլիգոսախարիդներ
խմբագրելԴիսախարիդներ
խմբագրելԵրկու մոնոսախարիդներից կազմված գլիկոզիդները կոչվում են դիսախարիդներ։ Դիսախարիդների գլիկոզիդային կապի առաջացմանը մասնակցում են երկու OH-խմբեր՝ մոնոսախարիդներից մեկի կիսաացետալայինը և մյուսի՝ ցանկացածը։ Եթե երկրորդ հիդրօքսիլ խումբը սպիրտային է, ապա նման միացության մեջ պահպանվում է կիսաացետալային խումբը և այդպիսի դիսախարիդները կոչվում են վերականգնող (լակտոզ, մալթոզ, ցելոբիոզ) կամ մալթոզային տիպի։ Եթե գլիկոզիդային կապի առաջացմանը մասնակցում են երկու մոնոսախարիդների կիսաացետալային խմբերը, ապա այդպիսի դիսախարիդները կոչվում են տրեգալոզային կամ չվերականգնող տիպի։ Դիսախարիդների հիդրոլիզի արդյունքում առաջանում են մոնոսախարիդներ։
սախարոզ+H2O=D-գլյուկոպիրանոզ+D-ֆրուկտոֆուրանոզ
լակտոզ+H2O=D-գլյուկոպիրանոզ+D-գալակտոպիրանոզ
մալտոզ+H2O=D-գլյուկոպիրանոզ+D-գլյուկոպիրանոզ
ցելոբիոզ+H2O=D-գլյուկոպիրանոզ+D-գլյուկոպիրանոզ

Դիսախարիդները ըստ կառուցվածքի O-գլիկոզիդներ են, որոնցում գլիկոնային մասը նույնպես ներկայացված է մոնոսախարիդով։ Դիսախարիդների անվանակարգը կատարվում է համաձայն այն կանոնների, որոնք կիրառում են գլիկոզիդների համար.կապի առաջացմանը կիսաացետալային խումբ տրամադրող մոնոսախարիդը փոխում է իր վերջավորությունը «ոզիլ»-ի, իսկ սպիրտային խումբ տրամադրող մոնոսախարիդի վերջավորությունը չի փոխվում։
Դիսախարիդների անվան մեջ անպայման նշվում է երկու մոնոսախարիդների լրիվ կոնֆիգուրացիաները, օրինակ մալթոզի անվանումն է՝ α-D-գլյուկոպիրանոզիլ-(1→4)-α(β)-D-գլյուկոպիրանոզ։ Սախարոզի մոլեկուլի առաջացմանը մասնակցում են երկու մոնոսախարիդների կիսաացետալային խմբերը և նրա անվանումն է՝ α-D-գլյուկոպիրանոզիլ-(1→2)-β-D-ֆրուկտոֆուրանոզիդ։
Վերականգնող դիսախարիդները հանդես են գալիս տաուտամեր ձևերով և նրանց թարմ ջրային լուծույթները ենթարկվում են մուտառոտացիայի։ Մալթոզը առավել տարածված դիսախարիդներից է, առաջանում է օսլայի և գլիկոգենի ֆերմենտային հիդրոլիզի արդյունքում աղեստամոքսային տրակտի տարբեր հատվածներում գործող ֆերմենտների (թքի և ենթաստամոքսային գեղձի կողմից արտադրված α-ամիլազների) ազդեցության շնորհիվ։ Լուծույթներում միշտ առկա են մալթոզի բոլոր ձևերը, բայց գերակշռում են α- և β- անոմերների պիրանոզային ձևերը։ Շնորհիվ բաց ձևի գոյության, դիսախարիդները կարող են մոնոսախարիդների նման վերականգնել մետաղների իոնները։
Արդյունքում ալդեհիդային խումբը օքսիդանում է, վերածվելով կարբօքսիլ խմբի։ Առաջացած միացությունները կոչվում են բիոնաթթուներ։ Նման հատկություններ են ցուցաբերում կաթնաշաքարը՝ լակտոզը, որի լուծույթը նույնպես տաուտոմեր ձևերի խառնուրդ է։

Գլիկոզիդային կապը կարող է առաջանալ նաև C2,C3,C4,C6 դիրքերում գտնվող հիդրօքսիլ խմբերի հաշվին.օրինակ իզոմալթոզում գլիկոզիդային կապը գոյացել է առաջին մոլեկուլի C1 և երկրորդ մոլեկուլի C6 միջև։


Որոշ դեպքերում գլիկոզիդային կապի առաջացմանը մասնակցում են երկու մոնոսախարիդների կիսաացետալային խմբերը, այնպես, ինչպես դա տեղի է ունենում սախարոզում։
Սախարոզում չկա ազատ կիսաացետալային հիդրօքսիլ խումբ։ Նման օլիգոսախարիդները չունեն տաուտոմեր ձևեր, չեն ցուցաբերում վերականգնողական հատկություններ և կոչվում են չվերականգնող օլիգոսախարիդներ։ Նման կառուցվածք ունեցող դիսախարիդների թարմ ջրային լուծույթները չեն ենթարկվում մուտառոտացիայի։
Օլիգոսախարիդների քիմիական հատկությունները
խմբագրելԴիսախարիդները ենթարկվում են հիդրոլիզի։ Օրգանիզմում հիդրոլիզը կատարվում է գլիկոզիդազ ֆերմենտների ազդեցությամբ։ Սախարոզը (ինվերտային շաքար) սննդի ածխաջրերի հիմնական աղբյուրն է, հեշտությամբ հիդրոլիզվում է թքի ֆերմենտներով, առաջացնելով գլյուկոզ և ֆրուկտոզ։
C12+H2O→C6H12O6(գլյուկոզ)+C6H12O6(ֆրուկտոզ)
Սախարոզը ինվերտային շաքար է կոչվում, քանի որ նա օպտիկապես աջ պտտող շաքար է, իսկ հիդրոլիզի արդյունքում առաջացած միացությունների՝ գլյուկոզի և ֆրուկտոզի էկվիմոլյար խառնուրդի պտտման ուղղությունը ձախն է.այսինքն տեղի է ունենում պտտման ուղղության փոփոխություն (inversion-փոխել ուղղվածությունը)։ Բերված հիդրոլիզի սխեմայից երևում է, որ առաջացած մոնոսախարիդների պտտման անկյունները և ուղղությունները կտրականապես տարբերվում են և ֆրուկտոզի ունակությունը գերակշռում է։
Լակտոզի հիդրոլիզից առաջանում են գլյուկոզ և գալակտոզ, մալթոզի հիդրոլիզից՝ երկու գլյուկոզ։
Օլիգոսախարիդների քիմիական հատկությունները հիմնականում կրկնում են մոնոսախարիդների հատկությունները։ Լաբորատոր պայմաններում հիդրոլիզը կատարվում է թթվային միջավայրում, որի հետևանքով առաջանում է օլիգոսախարիդի կազմի մեջ մտնող մոնոսախարիդների խառնուրդ։
Պոլիսախարիդներ
խմբագրելՄոնոսախարիդների կամ նրանց ածանցյալների պոլիկոնդեսացման պրոդուկտները կոչվում են պոլիսախարիդներ։ Մոնոմերները այդ միացություններում կապված են միմյանց հետ գլիկոզիդային կապերով։ Պոլիսախարիդները կարող են լինել ճյուղավորված և չճյուղավորված։ Պոլիսախարիդների կազմի մեջ մտնում են զանազան մոնոսախարիդներ.D-գալակտոզ,D-գլյուկոզ,D-գլյուկուրոնաթթու և այլն։
Հոմոպոլիսախարիդներ
խմբագրելԵթե պոլիսախարիդները կազմված են միևնույն մոնոսախարիդների մնացորդներից, ապա նրանք կոչվում են հոմոպոլիսախարիդներ կամ գլիկաններ։ Գլիկանները կարող են լինել հեքսոզաններ և պենտոզաններ։ Կախված մոնոսախարիդի բնույթից տարբերում են գլյուկաններ (որոնք կազմված են միայն գլյուկոզի մնացորդներից),մանաններ, գալակտաններ և այլն։ Երկրագնդի վրա ամենատարածված օրգանական միացությունը ցելյուլոզն (բջջանյութ) է, որը կազմված է 1,4-գլիկոզիդային կապով կապված β-D-գլյուկոպիրանոզի մնացորդներից, ունի կարևոր կառուցվածքային, հենարանային ֆունկցիա բուսական աշխարհում և անհրաժեշտ է որպես սննդի բաղադրամաս կենդանական աշխարհի ներկայացուցիչների համար։

Բույսերում ածխաջրերի պահեստային կամ պաշարային ձևն է օսլան, որը կազմված է գծային կառուցվածք ունեցող ամիլոզից (10-20%), և ճյուղավորված ամիլոպեկտինից (80 %)։ Ամիլոզը կազմված է α-1,4-գլիկոզիդային կապերով կապված D-գլյուկոպիրանոզի մնացորդներից։
Ամիլոպեկտինի ճյուղավորումը կատարվում է հիմնական շղթայի յուրաքանչյուր 20-25 մոնոսախարիդային մնացորդի մոտ 1,6-α-գլիկոզիդային կապերի առաջացմամբ։ Կենդանական օրգանիզմում ածխաջրերի պահեստային ձևը գլիկոգեն պոլիսախարիդն է (կենդանական օսլա), որը կառուցվածքով նման է ամիլոպեկտինին, սակայն ճյուղավորված է.կողմնային շղթաները կարող են գոյանալ յուրաքանչյուր 6-րդ, 8-րդ մոնոսախարիդի մոտ։ Մոնոսախարիդները գլիկոգենում կապված են միմյանց հետ հիմնականում α-1,4- և α-1,6 գլիկոզիդային կապերով։
Դեքստրան։ Դեքստրանները մանրէների հիմնական պոլիսախարիդներն են։ Այժմ մշակված են դեքստրանների արդյունաբերական ստացման եղանակներ միկրոբիոլոգիական սինթեզի ճանապարհով։ Դեքստրանները կազմված են α-D-գլյուկոպիրանոզային մնացորդներից.մոլեկուլները խիստ ճյուղավորված են։ Կապի հիմնական ձևն է α(1→6), իսկ ճյուղավորման հատվածներում α(1→4), α(1→3), α(1→2)։
Դեքստրանները օգտագործում են արյան փոխարինիչների պատրաստման համար (պոլիգլյուկին)։ Ատամների վրա զարգացող մանրէները նույնպես արտադրում են դեքստրան, որը հայտնաբերվում է ատամների վրա գոյացող նստվածքի կազմում։

Մեկ այլ պոլիսախարիդ, որը մոնոմերն է D-ֆրուկտոզի մնացորդը, կոչվում է ինսուլին։ Ինսուլինը սոխի, սխտորի և մի շարք այլ մթերքների պահեստային պոլիսախարիդն է և օգտագործվում է ֆրուկտոզի արտադրության համար։
Հետերոպոլիսախարիդներ
խմբագրելԿենդանի օրգանիզմներում կարևոր դեր ունեն հետերոպոլիսախարիդները, որոնք կազմված են տարբեր մոնոսախարիդների մնացորդներից։ Հետերոպոլիսախարիդները հիմնականում կազմված են դիսախարիդային միավորներից։ Առավել մեծ նշանակություն ունեն հիալուրոնաթթուն, հեպարինը, խոնդրոիտինսուլֆատները և այլն։ Շարակցական հյուսվածքի հիմնական պոլիսախարիդն է հիալուրոնաթթուն, որը առաջին անգամ անջատվել է ապակյա մարմնից և սինովիալ հեղուկից, իսկ հետագայում հայտնաբերվել է մաշկում, պորտալարում, հեմոլիտիկ ստրեպտոկոկկում և այլն։ Հիալուրոնաթթվի հիդրոլիզից առաջանում են D-գլյուկուրոնաթթու,D-գլյուկոզամին և քացախաթթու.մոլեկուլն ունի գծային կառուցվածք, ճյուղավորված չէ, մոլեկուլային զանգվածը տատանվում է երեքից ութ միլիոնի սահմաններում։ Հյուսվածքներում հանդիպում է ինչպես ազատ, այնպես էլ սպիտակուցների հետ (աղանման) կապված։

Հեպարին։ Հեպարինը արյան հակամակարդիչ (հակակոագուլյանտ) գործոնն է, հայտնաբերվել է նաև լյարդում, փայծաղում, թոքերում, վահանագեղձում։ Հեպարինի կրկնվող օղակը բաղկացած է D-գլյուկուրոնաթթվից և D-գլյուկոզամինից, որում ամինախումբը և հիդրօքսիլ խմբերից որևէ մեկը կապված են ծծմբական թթվի մնացորդի հետ։

Խոնդրոիտինսուլֆատները զանազան հյուսվածքներում ներկայացված են իրենց երեք հիմնական ձևերով՝ A,B,C:Խոնդրոիտինսուլֆատ B-ն (որը հայտնի է նաև որպես β-հեպարին) և ավելի ուշ մաշկից անջատված դերմատանսուլֆատը պոլիսախարիդներ են, որոնց կրկնվող միավորը կազմված է L-իդուրոնաթթվի և N-ացետիլգալակտոզամինսուլֆատից։

Ամինագլիկանների շարքին է պատկանում նաև կերատանսուլֆատը, որի կրկնվող միավորը կազմված է β-D-գալակտոզի և N-ացետիլ-β-D-գլյուկոզամին մնացորդներից։
Մուկոպոլիսախարիդները կարևոր նշանակություն ունեն նաև մանրէների նյութափոխանակության և գոյատևման համար։ Օրինակ, պնևմոկոկկեր II և III պատիճներում հայտնաբերվել են սպեցիֆիկ պոլիսախարիդներ (ալդոբիուրոնաթթուներ),որոնք օժտված են անտիգենային հատկություններով։
Գլիկոպրոտեիններ
խմբագրելՕլիգո- և պոլիսախարիդները հաճախ են մտնում խառը բիոպոլիմերների կազմի մեջ, որոնցից կարևոր կենսաբանական դեր ունեն գլիկոպրոտեինները։ Այդ միացություններում սպիտակուցները սովորաբար կովալենտ կապով կապված են ածխաջրատային շղթաների հետ։ Գլիկոպրոտեինների խմբին են պատկանում իմունոգլոբուլինները և արյան խմբային նյութերը։ Ըստ կառուցվածքի դրանք բիոպոլիմերներ են, որտեղ սպիտակուցային շղթային միացած են բազմաթիվ ածխաջրային շղթաներ, հիշեցնելով փորձանոթներ լվացող խոզանակներ։ Ածխաջրային մասի զանգվածը կազմում է ընդհանուրի 80 %:Օլիգոսախարիդային հատվածների համար բնորոշ է վերականգնող ծայրերում մոնոսախարիդների (սովորաբար 3-5) որոշակի հաջորդականությունը։ Այդ ծայրերը կոչվում են անտիգենային դետերմինանտներ և պայմանավորում են արյան խմբային սպեցիֆիկությունը։ Դետերմինանտը փոխվելու դեպքում արյան խումբը նույնպես կփոխվի։ Արյան A-խմբի անտիգենային դետերմինանտն է N-ացետիլգալակտոզամինը,B-խմբինը՝ D-գալակտոզը,O-խմբինը՝ Լ-ֆուկոզը։
70-ական թվականներին, մշակելով B(III)-խմբի արյան էրիթրոցիտները գալակտոզիդազ ֆերմենտով, որը հիդրոլիզում է գալակտոզի հետ առաջացած գլիկոզիդային կապը, ստացել են O(I)-խմբի դետերմինանտները, այսինքն արյան III-խմբի էրիթրոցիտները վերածվել են I-խմբի էրիթրոցիտների։
Խառը բիոպոլիմերներում ածխաջրային հատվածը սպիտակուցի շղթայի հետ կապվում է երկու հնարավոր եղանակներով՝ C-N-գլիկոզիդային և C-O-գլիկոզիդային կապերով։ Սովորաբար սպիտակուցային հատվածը գտնվում է բջջի թաղանթում, իսկ ածխաջրայինը սուզված է շրջապատող հեղուկ միջավայրի մեջ, ապահովելով բջջի և միջավայրի կապը։
Մանրէների բջջաթաղանթը, ի տարբերություն բարձրագույն օրգանիզմների, ունի պաշտպանական ծածկ՝ պատ։ Մանրէների բջջապատը կառուցված է ածխաջրերից և պեպտիդներից կազմված միավորներից։
Թքի հակաբակտերիալ ակտիվությունը պայմանավորված է նրանով, որ լիզոցիմի (ֆերմենտի) հիդրոլիտիկ ազդեցության շնորհիվ քայքայվում են պոլիսախարիդային շղթայի գլիկոզիդային կապերը, այսինքն քայքայվում են մանրէների պատերը և դրանում է կայանում լիզոցիմի ազդեցության քիմիական մեխանիզմը։
Մանրէների բջջապատի սինթեզի վերջին փուլում գլիկոպրոտեինները փոխազդում են միմյանց հետ, առաջացնելով եռաչափ ցանց։ Նշված պրոցեսը կատարվում է ֆերմենտների ազդեցությամբ, որի արդյունքում առաջանում են կապեր՝ մի շղթայի D-Ալա մնացորդի և մյուս շղթայի Գլի-մնացորդի միջև։
Պենիցիլինի հակաբակտերիալ ազդեցության մեխանիզմը տարբերվում է լիզոցիմի ազդման մեխանիզմից.պենիցիլինի կոնֆիգուրացիան նման է պեպտիդգլիկանի Ալա-Ալա-ծայրային կառուցվածքին և նա կարող է զբաղեցնել տրանսպեպտիդազ ֆերմենտի (բջջաթաղանթի սինթեզը ապահովող) ակտիվ կենտրոնը, առաջացնելով ամուր, չքանդվող կապ ֆերմենտի և պենիցիլինի β-լակտամային օղակի կարբոնիլ խմբի միջև։ Այս փոխազդեցության հետևանքով խանգարվում է բջջի պատի սինթեզը, ամբողջականությունը, ձևավորումը և բջջի պարունակությունը արտահոսում է՝ արդյունքում բջիջը մահանում է։ Բարձրագույն օրգանիզմները նման բջջապատեր չունեն, հետևաբար պենիցիլինը տոքսիկ է միայն մանրէների համար։
Փոխանակության խանգարումներ
խմբագրելԱծխաջրերի պակասի հետևանքով արյան մեջ կուտակվում են ճարպաթթուների ոչ լրիվ օքսիդացման արգասիքները՝ այսպես կոչված «կետոնային մարմնիկները», որոնք տեղաշարժելով արյան ռեակցիան դեպի թթվային կողմ, խախտում են շատ ֆերմնետային համակարգերի գործառույթներ, ինչը նկատվում է շաքարային դիաբետով հիվանդների, թմրամոլների, հարբեցողների, տոկսիկոմանների մոտ։
Տարածված ածխաջրեր
խմբագրել
Գլյուկոզի ալֆա և բետա տարաձևերը (անոմերները), որոնք միմյանցից տարբերվում են 5-րդ ածխածնի ատոմին միացած -OH խմբի դիրքով (կարմիր կամ կանաչ)[7]
Միաշաքարներ
խմբագրելՀեքսոզներ
խմբագրելԱռավել տարածված միաշաքարներից՝ հեքսոզներից (մոլեկուլում 6 ածխածնի ատոմ պարունակող ածխաջրեր) են մանոզը, գալակտոզը գլյուկոզը։
Գլյուկոզ
խմբագրելԲնության մեջ առավել տարածված միաշաքարը խաղողաշաքարն է։ Ազատ վիճակում պարունակվում է քաղցր մրգերում, մարդու և կաթնասուն կենդանիների արյան պարտադիր բաղադրիչներից է։
Մանոզ
խմբագրելՄանոզը կարող է հանդիպել ազատ վիճակում, բայց առավել հաճախ այլ միաշաքարների հետ միացած գտնվում է երկար, բազմաշաքարային շղթաներում կամ գլիկոպրոտեիդների կազմության մեջ։
Գալակտոզ
խմբագրելԳալակտոզն ազատ վիճակում չի հանդիպում։ Այն խաղողաշաքարի հետ մտնում է կաթնաշաքարի, ինչպես նաև շատ բազմաշաքարների և գլիկոպրոտեիդների բաղադրության մեջ։ Գալակտոզի փոխանակության խանգարումը, օրգանիզմի կողմից այն չյուրացվելը հանգեցնում են ծանր ժառանգական հիվանդության՝ գալակտոզարյունության (գալակտոզեմիայի)։ Երեխան, սնվելով մայրական կաթով, ստանում է մեծ քանակությամբ գալակտոզ, որի օգտագործման խանգարման դեպքում նկատվում է քաշի աստիճանական անկում, ֆիզիկական և մտավոր թերզարգացում, ոսպնյակի մթագնում, լյարդի մեծացում՝ որոշ դեպքերում՝ ցիռոզ։ Հիվանդությունը կրծքի երեխաների համար կարոդ է մահացու լինել։ Գալակտոզարյունությունն ուղեկցող շատ ախտաբանական փոփոխություններ հետևանք են հյուսվածքներում և օրգաններում գալակտոզի փոխանակության արդյունքում թունավոր նյութերի կուտակման։ Ժամանակին ախտորոշելիս և սննդից կաթնաշաքարն ու գալակտոզը լրիվ հեռացնելիս երեխայի աճն ու զարգացումը բնականոն են ընթանում։
Հեպտոզներ
խմբագրել
Ամենակարևոր պենտոզներից (մոլեկուլում 5 ածխածնի ատոմ պարունակող ածխաջրերից) են ռիբոզը և դեզօքսիռիբոզը, որոնք մտնում են ռիբոնուկլեինաթթուների և դեզօքսիռիբոնուկլեինաթթուներիկազմության մեջ։ Ներկայումս հայտնի է մոտ 70 միաշաքար, որոնցից 20-ը հայտնաբերվել է բնության մեջ, մնացածը սինթեզվել արհեստական եղանակով։
Ֆրուկտոզ
խմբագրելՊտղաշաքարն ազատ վիճակում պարունակվում է մեղրում, որոշ մրգերում։ Խաղողաշաքարի հետ այն առաջացնում է սննդի համար առավել էական նշանակություն ունեցող եղեգնաշաքար։
Երկշաքարներ
խմբագրել
Երկշաքարներից կարևոր նշանակություն ունեն ածիկաշաքարը, կաթնաշաքարը և եղեգնաշաքարը։
Ածիկաշաքար
խմբագրելԱծիկաշաքարը կազմված է խաղողաշաքարի 2 մնացորդից։ Առաջանում է բույսերի և կենդանիների հիմնական պահեստային ածխաջրերի՝ օսլայի և գլիկոգենի մասնակի՝ հիդրոլիզային ճեղքման ժամանակ և ունի կարևոր սննդային նշանակություն։ Որոշ մարդիկ տառապում են ածիկաշաքարի և իզոածիկաշաքարի նկատմամբ բնածին անհանդուրժողականությամբ, որն աղիքահյութում դրանք ճեղքող ֆերմենտների բացակայության հետևանք է։
Կաթնաշաքար
խմբագրելԿաթնաշաքարը կազմված է գալակտոզի և խաղողաշաքարի մնացորդներից։ Մեծ քանակությամբ պարունակվում է կաթնասունների կաթում, մայրական կաթում՝ 5,5-8,4 %, կովի կաթում՝ 4-5 %: Կրծքի երեխաների համար սննդի կարևոր բաղադրամաս է։
Աղիքահյութում կաթնաշաքար ճեղքող ֆերմենտի (լակտազ) բնածին բացակայության դեպքում առաջանում է հիվանդություն, որը շատ վաղ է դրսևորվում և ուղեկցվում է փսխումով, լուծով, փորափքանքով, ջրազրկմամբ, աստիճան, նիհարումով։ Մեզի հետ հեռանում են զգալի քանակությամբ կաթնաշաքար և շատ ամինաթթուներ։ Բուժման համար սննդի կաթնաշաքարը պետք է փոխարինել եղեգնաշաքարով, խաղողաշաքարով և այլ շաքարներով։
Եղեգնաշաքար
խմբագրելԵղեգնաշաքարը կազմված է խաղողաշաքարի և պտղաշաքարի մնացորդներից։ Լայնորեն տարածված է բուսական աշխարհում և հիմնական սննդային ածաջրերից է։ Սննդի մեջ օգտագործվում է շաքարի ճակնդեղից և եղեգից ստացված շաքարը։
Աղիքահյութում վերջինս ճեղքոդ ֆերմենտի՝ սախարազի բացակայության պատճառով առաջանում է հիվանդություն, որը դրսևորվում է եղեգնաշաքարի նկատմամբ օրգանիզմի ժառանգական անհանդուրժողականությամբ։ Երեխաների մոտ այն առաջանում է խառը սնուցման անցնելիս։ Նկատվում է լուծ, երեխան նիհարում է, եղեգնաշաքարը չի յուրացվում և դուրս է գալիս օրգանիզմից։ Բուժման նպատակով օրաբաժնից պետք է հանել եղեգնաշաքարը կամ սննդի հետ տալ համապատասխան ֆերմենտների պատրաստուկներ։
Բազմաշաքարներ
խմբագրել

Բազմաշաքարները ածխաջրերի բարձրամոլեկուլային բնական միացություններ են՝ կազմված մեծ քանակությամբ միաշաքարների օղակներից։ Կատարում են կառուցվածքային և սննդային ֆունկցիա։ Բնության մեջ առավել տարածված են թաղանթանյութը, օսլան և գլիկոգենը։
Թաղանթանյութ
խմբագրելԹաղանթանյութը (բջջանյութ, ցելյուլոզ) բուսական հյուսվածքների հիմնական կառուցվածքային բաղադրամասն է։ Պարունակվում է հիմնականում բուսական բջիջների պատերում։ Մոլեկուլը բաղկացած է ուղիղ, երկար կապերով միմյանց միացած խաղողաշաքարի մնացորդներից։ Վատ մարսվելիության շնորհիվ այն խթանում է մարդու աղիքների գալարակծկանքը, կարգավորում դրանց գործունեությունը։
Օսլա
խմբագրելՕսլան բույսերի հիմնական պահեստային սննդանյութն է։ Այդ կարևորագույն սննդային բազմաշաքարը մեծ քանակությամբ պարունակվում է կարտոֆիլի պալարներում, հացահատիկներում, մրգերում։
Գլիկոգեն
խմբագրելԳլիկոգենը կամ կենդանական օսլան մարդու և կենդանիների կարևոր պահեստային բազմաշաքար է։ Օրգանիզմում այն հիմնականում կուտակվում է լյարդում (մինչև 20 %), մկաններում (4 %)։ Բազմաշաքարների փոխանակության խանգարման հետևանքով առաջանում են ծանր ժառանգական հիվանդություններ, որոնք չեն բուժվում հատուկ սննդակարգով։ Դրանցից են գլիկոգենոզները (բնածին), որոնք բնորոշվում են սարդում, սրտամկանում, կմախքային մկաններում, երիկամներում, փայծաղում, թոքերում և այլ օրգաններում գլիկոգենի մեծ կուտակումներով։ Հիվանդության բնորոշ ախտանշան է արյան մեջ խաղողաշաքարի ցածր կոնցենտրացիան (թերշաքարարյունություն) այն դեպքում, Երբ գլիկոգենի պարունակությունը հյուսվածքներում մեծ է, որը չի ենթարկվում ադրենալինի և գլյուկագոնի ազդեցությանը։ Կուտակվելով բջիջներում՛ գլիկոգենն առաջացնում է դրանց կազմափոխություն և մահ։ Գլիկոգենոզի դեպքում նկատվում է մկանային թուլություն, աճի կասեցում, ճարպակալում, ախտահարվում են լյարդը և երիկամները։ Հիվանդությունները կապված են ածխաջրային փոխանակությանը մասնակցող մի շարք ֆերմենտների բացակայության հետ, ընդ որում՝ յուրաքանչյուր տեսակի գլիկոգենոզի համար բնորոշ է որոշակի ֆերմենտի անբավարարությունը, թեպետ կա նաև խառը տիպի հիվանդություն։
Մուկոբազմաշաքարներ
խմբագրելՄարդու և կաթնասուն կենդանիների շարակցական հյուսվածքի ձևավորման և բնականոն գործունեության համար կարևոր նշանակություն ունեն մուկոբազմաշաքարները, օրինակ՝ հիալուրոնաթթուն, քոնդրիոտինսուլֆատները։
Ածխաջրերի փոխանակություն
խմբագրել
Բարդ ածխաջրեր, սննդի հետ թափանցելով մարդու օրգանիզմ, մարսողության համակարգի բազմաթիվ ֆերմենտներով ճեղքվում են մինչև միաշաքարների, որոնք աղիքներից ներծծվում են արյան մեջ, լուծված վիճակում անցնում են բարդ և հատուկ ֆերմենտների օգնությամբ փոխարկվում խաղողաշաքարի։ Վերջինիս մի մասը վերածվում է գլիկոգենի, մյուսը՝ արյան հոսքով տարվում է օրգաններ ու հյուսվածքներ և օգտագործվում գլիկոգենի, ճարպերի ու փոխարինելի ամինաթթուների սինթեզի համար։
Գլյուկոզի փոխանակություն
խմբագրելԳլյուկոզի կոնցենտրացիայի պահպանումը արյան մեջ
խմբագրելԳլյուկոզի հաստատուն կոնցենտրացիան արյան մեջ պահպանվում է չափազանց բարդ մեխանիզմի օգնությամբ, որին մասնակցում են մեծ քանակությամբ ֆերմենտներ, հորմոններ (ինսուլին, գլյուկագոն) և նյարդային համակարգը։ Սննդի հետ շատ ածխաջրեր օգտագործելիս խաղողաշաքարի բնականոն կոնցենտրացիան արյան մեջ և հյուսվածքներում հնարավոր է պահպանել ցարդի ու հատկապես մկանների բջիջների կողմից՝ դրանից պահեստային բազմաշաքարը՝ գլիկոգենը, սինթեզելու շնորհիվ։ Վերջինս անհրաժեշտության դեպքում մի շարք բարձրմասնագիտացված ֆերմենտների օգնությամբ կարող է նորից փոխարկվել խաղողաշաքարի, որը կարևոր նշանակություն ունի բուսական և կենդանակ բջիջներում ընթացող նյութափոխանակության մեջ, որպես էներգիայի աղբյուր։
Գլյուկոզի օքսիդացում
խմբագրելՖերմենտային ռեակցիաների բարդ շղթայի հետևանքով խաղողաշաքարն օքսիդանում է՝ առաջացնելով ածխաթթու գազ և ջուր, որն ուղեկցվում է ադենոզինեռֆոսֆորաթթվի (ԱԵՖ) առաջացմամբ։ Վերջինիս մոլեկուլի ֆոսֆոր-թթվածին քիմիական կապի էներգիան այնքան մեծ է, որ կարող է ապահովել էներգիայի ծախսով ընթացող շատ կենսաբանական գործընթացներ (սպիտակուցների, նուկլեինաթթուների, ճարպերի, բարդ ածխաջրերի կենսասինթեզ, մկանային կծկում և այլն)։ Խաղողաշաքարի փոխանակության միջանկյալ նյութերը մի շարք դեպքերում ընդհանուր են դառնում ամինաթթուների և ճարպաթթուների փոխանակության միջանկյալ նյութերի հետ։ Բացի այդ, որոշ ամինաթթուներից կարող է սինթեզվել խաղողաշաքար՝ ֆոսֆորային ածանցյալների ձևով, որը գլիկոգենի փոխանակության հետ միասին կարևոր նշանակություն ունի հյուսվածքներում և արյան մեջ խաղողաշաքարի հաստատուն կոնցենտրացիայի պահպանման գործում։ Այսպիսով՝ կենդանի օրգանիզմի բջիջներում սերտ կապ գոյություն ունի ածխաջրերի, սպիտակուցների ու ճարպերի փոխանակության միջև։ Խաղողաշաքարը մասնակցում է նաև այլ միաշաքարների առաջացման գործընթացներին։
Փոխանակության խանգարումներ
խմբագրել
Հիպերգլիկեմիա
խմբագրելՄեծ քանակությամբ խաղողաշաքարի անցումն աղիքներից արյան մեջ հանգեցնում է սննդային գերշաքարարյունության, որն արդյունք է ածխաջրերով հարուստ սննդի ընդունման։ Խիստ բարձր սննդային գերշաքարարյունության դեպքում խաղողաշաքարը հեռացվում է մեզի հետ (շաքարամիզություն), որն առողջության համար վտանգավոր չէ։
Հուզումների ժամանակ գլխուղեղի կեղևում առաջացած դրդումը տարածվում է երկարավուն ուղեղ, որտեղից սիմպաթիկ նյարդի ճյուղերով՝ մակերիկամների միջուկային նյութին։ Դա հանգեցնում է ադրենալինի արտազատման ուժեղացմանը և նպաստում լյարդում գլիկոգենի ճեղքմանը մինչև խաղողաշաքարի, որն ավելցուկային քանակությամբ անցնում է արյան մեջ՝ առաջացնելով հոգեկան կամ հուզական գերշաքարարյունություն։
Թունավորումների, գլխուղեղի վնասվածքների, որոշ վարակիչ հիվանդությունների դեպքում գերշաքարարյունությունը կարող է առաջանալ որպես կենար, նյարդային համակարգի գրգռման հետևանք։ Խաղողաշաքարի պարունակությունն արյան մեջ մեծանում է բարձր ջերմաստիճանի, ջոկային վիճակների, ուղեղապատյանաբորբերի, գլխուղեղի բորբոքումների ժամանակ։
Ենթաստամոքսային գեղձի ներզատիչ հյուսվածքի թերֆունկցիայի դեպքում ինսուլինի առաջացման կտրուկ նվազման հետևանքով փոխվում է ինսուլին-ադրենալին փոխհարաբերությունը, առաջանում է ինսուլինային գերշաքարարյունություն։
Հիպոգլիկեմիա
խմբագրելՍննդային գերշաքարարյունությունը 3-5 ժ հետո փոխարինվում է ուժեղ թերշաքարարյունությամբ, որը կապված է ինսուլինի արտազատման ուժեղացման հետ։ Թերշաքարարյունությունը կարող է առաջանալ ծանր և երկարատև ֆիզիկական աշխատանքի, ենթաստամոքսային գեղձի ներզատիչ հյուսվածքի ախտահարման, երիկամների հիվանդությունների, բրոնզախտի, լորձայտուցի և այլ դեպքերում։ Հատուկ ձևի թերշաքարարյունություն է առաջանում բուժիչ նպատակով ինսուլինի չափաքանակը մեծացնելիս։ Ինքնաբեր թերշաքարարյունություն նկատվում է նյարդային համակարգի հիվանդությունների և հոգեկան խանգարումների դեպքերում։ Հյուսվածքների, առանձնապես գլխուղեղի, ածխաջրային (խաղողաշաքարային) քաղցը կարող է հանգեցնել կոմայի։ Օրգանիզմում խաղողաշաքարի անբավարարության վաղ դրսևորումներից են քաղցի զգացումը, քրտնոտությունը, որոնք վերանում են սննդային շաքարի, հացի, մրգահյութերի ընդունումով։
Ժառանգական խանգարումներ
խմբագրելՀայտնի են ածխաջրերի փոխանակության խախտումների հետ կապված մի շարք ժառանգական խանգարումներ, որոնք բնորոշվում են տարբեր հյուսվածքների բջիջներում ածխաջրերի բարդ միացությունների կուտակումներով։ Կուտակվող նյութերի կազմությունից կախված՝ այդ հիվանդությունները լինում են մուկոբազմասախարիդոզներ, գլիկոլիպիդոզներ և գլիկոպրոտեիդոզներ, որոնց առաջացումը պայմանավորված է մուկոբազմաշաքարների, գլիկոլիպիդների կամ գլիկոպրոտեիդների ճեղքմանը մասնակցող համապատասխան ֆերմենտի բացակայությամբ կամ ակտիվության կտրուկ փոքրացմամբ։
Սննդանյութեր
խմբագրել
Ածխաջրերը սննդի մեջ էներգիական արժեքով ածխաջրեր հավասարարժեք են սպիտակուցներին։ 1 գ ածխաջրի այրումից օրգանիզմում անջատվում է միջինը 4,1 կկալ էներգիա։ Մարդու սննդի մեջ ածխաջրերի քանակը սովորաբար 4 անգամ գերազանցում է սպիտակուցների և ճարպերի պարունակությանը։ Օրգանիզմում ածխաջրերի սահմանափակ կուտակվելու հատկությունը նպաստում է դրանց ավելցուկային քանակների հեշտորեն ճարպի փոխարկվելուն, որը կուտակվում է ճարպային պահեստարանում (դեպոյում)։ Սննդի մեջ ածխաջրերի ավելցուկից առաջանում է ավելորդ քաշ և գիրություն։
Ածխաջրերի պարունակությունը սննդի օրաբաժնում պետք է հաշվեկշռված լինի սպիտակուցների և ճարպերի հետ։ Դրանց կտրուկ սահմանափակումը, առավել ևս լրիվ բացակայությունը, խանգարում է բնականոն սնուցումը և կարող է հանգեցնել օրգանիզմի կենսագործունեության էական փոփոխությունների։ Միայն բուժիչ նպատակով որոշ հիվանդությունների (շաքարախտ, ճարպակալում և այլն) դեպքում բժիշկը կարող է նշանակել ածխաջրերի ցածր պարունակությամբ սննդակարգ։
Ածխաջրեր սննդային օրաբաժնի հիմնական մասն են և ապահովում են օրգանիզմին անհրաժեշտ օրական կալորիականության կեսից ավելին։ Դրանց 1 օրվա ծախսը կազմում է 350-500 գ, որոնց նկատմամբ պահանջը բավարարվում է բուսական մթերքների (հացահատիկ, կարտոֆիլ և այլն), մասամբ էլ շաքարի հաշվին։ Կենդանական մթերքը, որպես ածխաջրերի աղբյուր, չնչին նշանակություն ունի։ Բայց վաղ մանկական տարիքում, երբ կաթը հիմնական սնունդն է, այն ածաջրերի կարևոր աղբյուր է և կարող է գրեթե լիովին ապահովել օրգանիզմի պահանջը։ Նույնը վերաբերում է նաև մեծահասակներին ածխաջրերով ապահովմանը։
Խաղողաշաքարի կարևոր հատկությունը հեշտ յուրացվելիությունն է և օրգանիզմի կողմից արագ օգտագործումը գլիկոգենի առաջացման համար, որն անհրաժեշտ է աշխատող մկանների, այդ թվում սրտամկանի սնուցման, արյան մեջ շաքարի բնականոն կոնցենտրացիան պահպանելու համար։ Խաղողաշաքարը սննդի լավագույն միջոց է հետվիրահատ. շրջանում, ինչպես նաև ծանր և թույլ հիվանդների համար։ Պտղաշաքարը մոտ 2 անգամ քաղցր է սննդային շաքարից և 3 անգամ՝ խաղողաշաքարից։ Այն առավել գործածական է մտավոր աշխատանք կատարելիս, ինչպես նաև հասուն և տարեց մարդկանց, աթերոսկլերոզով հիվանդների համար՝ ճարպային և խոլեստերինի փոխանակության խանգարումների դեպքում։
Պտղաշաքարը բարերար ազդեցություն է ունենում աղիքային օգտակար միկրոֆլորայի վրա։ Պտղաշաքարի և խաղողաշաքարի հիմն աղբյուր են մրգերը, հատապտուղները, ինչպես նաև մեղրը, որում խաղողաշաքարի պարունակությունը կազմում է 25-37 %, իսկ պտղաշաքարինը՝ 39-40 %: Սախարոզը շաքարի ձևով առավել օգտագործվող երկշաքար է։ Չնայած սննդային բարձր հատկություններին՝ երբեմն մտավոր աշխատանքի և քիչ շարժուն կենսակերպի, ճարպակալման հակում ունենալու դեպքերում, ինչպես նաև ծեր տարիքում մեծ քանակությամբ շաքարի օգտագործումը ցանկալի չէ։ Դրա ավելցուկը նպաստում է ճարպակալմանը և աթերոսկլերոզի առաջացմանը, որոշ տվյալներով՝ բարձրացնում է արյան մեջ խոլեստերինի պարունակությունը։
Սննդային օրաբաժնում օսլան կազմում է օգտագործվող ածխաջրերի ընդհանուր քանակի մոտ 80 %-ը, որի բարձր պարունակությամբ է մեծ չափով պայմանավորված հատիկային մթերքների, լոբազգիների և կարտոֆիլի սննդային արժեքը։ Օսլայի ճեղքման հաշվին է բավարարվում օրգանիզմի շաքարի, մասնավորապես՝ խաղողաշաքարի պահանջը։
Թաղանթանյութի և պեկտինային նյութերի հիմնական հատկությունը ջրային լուծույթում թթվի և շաքարի ներկայությամբ դոնդողանման կոլոիդային զանգված առաջացնելու ունակությունն է։ Հաստատվել է պեկտինային նյութերի նշանակությունն առողջ մարդու սննդի մեջ, ինչպես նաև դրանց օգտագործման հնարավորությունը ստամոքսաղիքային համակարգի որոշ հիվանդությունների, այրվածքների և այլ դեպքերում։ Հայտնի է բջջանյութի ածխաջրերի դերն աղիքների գալարակծկանքի խթանման գործում։ Բջջանյութը նպաստում է նաև օրգանիզմից խոլեստերինի հեռացմանը, աղիքային ֆլորայի բնականոնացմանը։
Գրականություն
խմբագրել- Бендер М., Бергерон Р., Комияма М. Биоорганическая химия ферментативного катализа. Пер. с англ. - М.: Мир, 1987. - 352 с.
- Дюга Г., Пенни К. Биоорганическая химия. - М.: Мир, 1983.
- Тюкавкина Н. А., Бауков Ю. И. Биоорганическая химия. - М.: Медицина, 1991.
Ծանոթագրություններ
խմբագրել- ↑ Н. А. АБАКУМОВА, Н. Н. БЫКОВА. 9. Углеводы // Органическая химия и основы биохимии. Часть 1. — Тамбов: ГОУ ВПО ТГТУ, 2010. — ISBN 978-5-8265-0922-7
- ↑ Eldra Pearl Solomon, Linda R. Berg, Diana W. Martin; Cengage Learning (2004). Biology. books.google.am. էջ 52. ISBN 978-0534278281.
{{cite book}}: CS1 սպաս․ բազմաթիվ անուններ: authors list (link) - ↑ Flitsch, Sabine L.; Ulijn, Rein V (2003). «Sugars tied to the spot». Nature. 421 (6920): 219–20. doi:10.1038/421219a. PMID 12529622.
- ↑ Maton, Anthea; Jean Hopkins; Charles William McLaughlin; Susan Johnson; Maryanna Quon Warner; David LaHart; Jill D. Wright (1993). Human Biology and Health. Englewood Cliffs, New Jersey, USA: Prentice Hall. էջեր 52–59. ISBN 0-13-981176-1.
- ↑ Campbell, Neil A.; Brad Williamson; Robin J. Heyden (2006). Biology: Exploring Life. Boston, Massachusetts: Pearson Prentice Hall. ISBN 0-13-250882-6.
- ↑ Pigman, Ward; Horton, D. (1972). «Chapter 1: Stereochemistry of the Monosaccharides». In Pigman and Horton (ed.). The Carbohydrates: Chemistry and Biochemistry Vol 1A (2nd ed.). San Diego: Academic Press. էջեր 1–67.
- ↑ http://www.ncbi.nlm.nih.gov/books/NBK1955/#_ch2_s4_
Արտաքին հղումներ
խմբագրել- «Ածխաջրեր» (ռուսերեն). Արխիվացված է օրիգինալից 2001 թ․ հուլիսի 25-ին. Վերցված է 2009 թ․ հունիսի 1-ին.
Տես նաև
խմբագրել| Վիքիպահեստն ունի նյութեր, որոնք վերաբերում են «Ածխաջուր» հոդվածին։ |
| Այս հոդվածի կամ նրա բաժնի որոշակի հատվածի սկզբնական կամ ներկայիս տարբերակը վերցված է Քրիեյթիվ Քոմմոնս Նշում–Համանման տարածում 3.0 (Creative Commons BY-SA 3.0) ազատ թույլատրագրով թողարկված Հայկական սովետական հանրագիտարանից (հ․ 1, էջ 209)։ |
| Այս հոդվածն ընտրվել է Հայերեն Վիքիպեդիայի օրվա հոդված: |